2. Anatomy of the Mediterranean
We cannot understand changes in any environment without
a basic grasp of the fundamental principles that govern that environments
physical, chemical, and biological characteristics. This chapter therefore
introduces in general terms the main processes ongoing in the Mediterranean
basin, as unraveled by oceanographers over especially the last 50 years.
The functioning of the basin can be likened to that of
an enormous organism. In fact as do many ocean basins the Mediterranean
breathes, taking in oxygen by exchange with the atmosphere at its surface,
and it has a distinct circulation system that moves the oxygen and other
chemical properties around the basin. Combined, these aspects affect the
plant and animal life in the basin, while this biological activity in turn
exerts its own influences on the distribution of chemical properties such
as nutrients and oxygen.
2.1. Lungs of the Ocean
Everywhere, oxygen concentrations in the oceans surface
waters are in an active process of exchange, or equilibration, with oxygen
concentrations in the atmosphere. This equilibration constitutes the only
significant route of oxygen into the oceans. Due to the equilibration,
oxygen levels in surface waters eventually reach what we call saturation,
meaning that for that waters particular temperature and salinity, the
concentration of oxygen is fixed not too high, not too low.
The entire surface layer, which amounts to the upper 150-200
meters or so that is stirred and mixed due to wind and wave action, thus
becomes well oxygenated. However, the oxygen is not efficiently passed
on to deeper layers. To understand why this is the case, we need to consider
the changes with depth in sea-water density (where density is weight per
litre).
In simplified terms, this problem can be portrayed by
assuming that the ocean is made up of discrete layers of water with different
characteristics. The surface layer has a lower density than the deeper
layer, which allows the surface layer to float on the deeper one. Its
like low-density oil floating on higher-density water, but with a smaller
density contrast. The density difference inhibits mixing between the two
layers, and so precludes an efficient exchange of properties (eg., oxygen)
between them. As a result, there is a very limited transfer into the deeper
layer of the oxygen that was taken up into the surface layer via equilibration
with the atmosphere.
Isolated from the atmosphere by the overlying surface
layer, the deep layer consequently would be seriously deprived of oxygen.
Still, many ocean basins, including the deep Mediterraean today, are quite
well oxygenated. This is caused by substantial oxygen transport into the
deep sea by localised, and often seasonal, formation of connections between
the deeper layer and the atmosphere. The specific locations where such
connections develop are described as deep-water formation areas. They
are the vital sites for oxygen transport into the deep sea, much the same
as our lungs are for oxygen transport into our bodies.
* * *
So what characteristics of sea water determine its density,
what happens in deep water formation areas, and how do these processes
operate in the Mediterranean?
The density of sea water depends on two properties: Temperature
and Salinity. Density decreases as temperature goes up, and increases as
temperature goes down (note
2). Hot air balloons provide a familiar illustration of temperatures
influence on density. As the burner heats the air within the balloon, its
density is lowered, causing the balloon to rise. When the burner is then
left off, the air in the balloon gradually cools by exchange with the atmophere,
its density increases, and the balloon gradually comes down. Besides temperature,
salinity also has a major effect on sea-water density. The more dissolved
salts per litre ie., the higher the salinity the higher the density.
At any given temperature, therefore, fresh water has a lower density than
sea water. Conversely, sea water that has undergone substantial evaporation
will have a higher density than normal sea water, because the evaporation
had removed pure water and left the salts behind.
Taking these two crucial influences on density into account,
we can predict that high densities in oceanic surface waters should be
found in poleward regions, where temperatures are very low. Highest values
should be expected in areas where mighty, saline, surface ocean currents
from the warm evaporative subtropics penetrate into such regions. In general
terms, this is exactly what we see. Because of the low temperatures, polar
to subpolar (high-latitude) regions show very high densities, but especially
high values occur between Scandinavia and Greenland, where salty waters
transported north by the Gulf Stream are subjected to high-latitude cooling.
As surface density values are increased, the contrast
between the surface and deeper layers fades. In particular places in the
world ocean, during particular seasonal conditions, this leads to the remarkable
condition that density becomes equal through the water column from the
surface to great depth. Nothing then stops violent mixing throughout the
water column and the consequent formation of new deep water. Thus, a connection
is established between the deep sea and the surface where equilibration
with the atmosphere takes place and the formation of new deep water pumps
fresh oxygen into the deep layers.
For the world ocean, the most important deep-water formation
areas are in the Labrador Sea and between Greenland and Scandinavia in
the northernmost Atlantic (North Atlantic Deep Water NADW), and in the
Weddell Sea to the east of the Antarctic Peninsula (AntArctic Bottom Water
AABW). Neither of those global deep-water masses can penetrate the Mediterranean,
however, because of a shallow sill (284 m) in the Strait of Gibraltar.
Why then do observations demonstrate that Mediterranean deep waters are
so well oxygenated? Does the basin perhaps have its own regions of deep-water
formation? The answer is Yes.
In the northern sector of the western Mediterranean (Gulf
of Lions), and in the northern basins of the eastern Mediterranean (Adriatic
and Aegean Seas), active deep-water formation takes place. The Gulf of
Lions deep water oxygenates the western Mediterranean between the Strait
of Gibraltar and the Strait of Sicily, while the Adriatic and Aegean deep
waters oxygenate the eastern Mediterranean. There is, however, a complication.
Between the surface layers and the deep waters, a distinct intermediate
water layer is recognised throughout the Mediterranean between 150/200
m and 600 m depth. In the next section, I will discuss the processes involved
in Mediterranean deep-water formation, including the vital role played
by the intermediate water.
2.2. Circulation
The deep circulation of the Mediterranean is driven
by a two-stage motor. In essence, surface waters in the Mediterranean have
their origin in the inflow of Atlantic waters through the Strait of Gibraltar.
Flowing eastwards through the Mediterranean, the surface waters are warmed
up by solar irradiation, and evaporation causes a steady increase in salinity.
As the inflowing waters, by now known as Modified Atlantic Water, arrive
in the easternmost Mediterranean, the salinities are very high indeed
they reach more than 39 salinity units, compared with about 36 in the original
Atlantic inflow, or with the global oceans average salinity of 34.5 units.
This high salinity in the easternmost Mediterranean would cause a high
density, if it were not compensated by very high temperatures, which in
summer reach 26ºC or more.
 Figure 2. Highly schematic representation of the two-stage deep ventilation
engine. Inflowing surface water is transformed into intermediate water
by a net salinity increase. The salt in the intermediate layer preconditions
the second stage, whose direct forcing is essentially thermal. The endresult
is the formation of deep water layer that is less saline, but colder, than
the intermediate water. Click on thumbnail for full-sized figure.
Figure 2. Highly schematic representation of the two-stage deep ventilation
engine. Inflowing surface water is transformed into intermediate water
by a net salinity increase. The salt in the intermediate layer preconditions
the second stage, whose direct forcing is essentially thermal. The endresult
is the formation of deep water layer that is less saline, but colder, than
the intermediate water. Click on thumbnail for full-sized figure.
Crucially, winter cooling reduces sea surface temperatures
in the general easternmost Mediterranean to around 17ºC, while a specific
area between Cyprus and Rhodes gets cooled that little bit extra by cold
winds from the Anatolian mountains. There, temperatures below 16ºC
are reached, closely appoximating those in the original inflow into the
basin. The density increase due to high salinity is now no longer compensated
by temperature however, and, as a consequence, deep-water formation takes
place in the area between Cyprus and Rhodes. This water does not sink all
the way to the bottom, but settles between 150/200 and 600 m depth we
call it Levantine Intermediate Water. As described, the formation of
this water mass is essentially driven by a salinity increase. The first
stage of the Mediterranean deep circulation motor, therefore, is salt-driven.
The Levantine Intermediate Water is found everywhere in
the Mediterranean. Spilling through the Strait of Sicily at depth, it is
also distinctly present in the western Mediterranean. With its high salinity,
it provides an enormous subsurface supply of salt to the entire basin that
is vital for the next stage of the Mediterranean deep circulation motor.
Along the northern margin of the Mediterranean, we observe
the coldest winter conditions in places influenced by cold polar/continental
air streams. These air flows are channelled over those areas by the mountain
ranges around the Mediterraneans northern margin. In the west, such channelling
through the Rhone Valley towards the Gulf of Lions gives rise to the Mistral
winds. Over the Adriatic Sea, similar conditions cause the Bora winds.
Again similar conditions occur over the Aegean Sea in winter (Vardar).
These cold and dry winds cause strong local cooling of the surface waters,
to temperatures of 11-13ºC. The resultant rise in density is kept
in check by the fact that salinities are not very high along the relatively
wet northern borderlands of the Mediterranean. Still, the cooling effects
do suffice to eliminate the density contrast between cool and relatively
fresh surface waters, and the underlying warmer but very salty Levantine
Intermediate Water. Mixing between those two water masses becomes possible.
Then, something extraordinary happens. Where mixing takes
place of two water masses of fundamentally different properties, but similar
density, the endproduct will have a higher density than the original components.
In our case, the endproducts are known as the Western Mediterranean Deep
Water (WMDW) from the Gulf of Lions, and the Eastern Mediterranean Deep
Water (EMDW) from the Adriatic and Aegean Seas.
The formation of EMDW and WMDW represents the second stage
of the deep circulation motor. The second stage, therefore, is dominantly
thermally driven by winter cooling. However, to function properly, it
requires the extra salt supplied by the first stage of the motor the
primarily salt-driven intermediate water formation.
With its two-stage deep-water formation, driven by net
evaporation and net cooling, the Mediterranean efficiently converts surface
water into subsurface water. A compensating inflow of new surface water
occurs from the Atlantic, through the Strait of Gibraltar. Meanwhile, the
active deep-water formation ensures that the deep basin remains well ventilated.
This process is so vigorous that it only needs about one century to produce
enough new deep water to replace the entire volume of Mediterranean deep
water. The thus replaced subsurface waters exit from the basin into the
Atlantic, through the Strait of Gibraltar. Because of the density differences,
the higher-density Mediterranean outflow (temperature ~13ºC, salinity
~38) takes place at depth, underneath the lower-density Atlantic inflow
(temperature ~16ºC, salinity ~36).
2.3. The Life that is Sustained
Generally speaking, plants form the base of the food
web. Landplants use water and mineral nutrients (especially phosphate and
nitrate) from the soil, and carbon from atmospheric CO2, and
combine these compounds to form new biomass (ie., to grow) via a chemical
reaction that requires energy from sunlight. During the process called
photosynthesis for combination/ creation with light oxygen is released.
Upon death, the biomass undergoes decomposition, a process that involves
microbes and that can either take place within an animals gut or out in
the open (eg., a rotting log). It goes under the technical name respiration,
and effectively reverses the photosynthetic process: the biomass is broken
down and the energy and nutrients that were used in its formation are released.
Hence, by eating grass, a cow indirectly consumes solar enery. The animal
stores this energy as sugars or fat.
In the oceans, the plants are represented by a rich diversity
of phytoplankton which translates as drifting plants. These microscopic
marine plants do exactly the same as their larger cousins on land. They
take up nutrients and carbon from the ambient sea water, and use solar
energy in photosysnthesis to form new biomass, and oxygen as a byproduct.
For photosynthesis, phytoplankton requires light. In the Mediterranean
Sea, light penetrates to a maximum depth of about 120 m. This restricts
the phytoplankton to the surface layer.
Phytoplankton forms the basis of the marine food web
it is therefore common practice to refer to phytoplankton organisms as
primary producers. By harnessing solar energy, phytoplankton forms the
vital start of energy flow to all animals grazing upon it, or predating
and scavenging upon other animals. Marine food webs are highly complex,
just like those on land, but in essence everything comes down to the transfer
of originally solar energy from the plants up though the food
web.
Since death and decomposition (respiration) are as common
in the sea as on land, most of the nutrients and carbon originally fixed
in biomass are eventually recycled back into the water column. In the illuminated
surface waters, however, dissolved nutrient concentrations remain low,
because any released nutrients get immediately taken up again for photosynthesis.
Below the light-penetration limit, in deeper waters, the story is different.
There is no photosynthesis at those depths, so that dissolved nutrients
are not being taken up again. As a consequence, nutrient concentrations
in deep-sea waters increase with time.
Oxygen concentrations in the water column follow the opposite
pattern to nutrients. High concentrations are found in the surface layer,
due to ongoing equilibration with the atmosphere. Below the surface layer,
however, oxygen input is limited to that supplied by the process of deep
water formation, while respiration of biomass continues, consuming oxygen.
As a result, oxygen levels drop steadily from the (near) saturation values
in newly-formed deep waters to progressively lower values in deep waters
of increasing age. Oxygen concentrations in the deep sea thus reflect a
precarious balance between ventilation by new deep water and oxygen consumption
by respiration.
Some nutrients are lost from the oceanic system, by becoming
trapped in the continuous process of sedimentation the formation of sea-floor
mud. This loss is offset by delivery of new nutrients to the sea via two
pathways.
One route is atmospheric. This route is particularly important
for nitrogen, which occurs in several gaseous forms in the atmosphere.
Nitrogen gas is especially abundant, constituting almost 80% of the air
we breathe. Gaseous nitrogen oxides are also common, and are enhanced by
the burning of fossil fuels and by decomposition of sewage. Specialised
algae can fix nitrogen from dissolved gases in surface waters, which equilibrate
with the atmosphere. Other, normally minor, atmospheric nutrient transports
involve dust and aerosols.
The other pathway for nutrients into the oceans, and the
dominant one in the case of phosphorus, is via rivers. Where the atmospheric
mechanism has fairly uniform influences over very large areas, the riverine
flux affects especially the coastal waters. Particularly strong impacts
are seen near river mouths. Today, rivers bring very high amounts of dissolved
nutrients into the sea, because they drain artificially fertilised lands,
and are contaminated with sewage. However, even before this massive anthropogenic
pollution, the dissolved nutrient influx via rivers was substantial. Even
in a system completely undisturbed by man, therefore, a massive increase
in river discharge into an enclosed sea would have fertilised that basin,
causing enhanced phytoplankton production. The main impact would, however,
always be rather limited to the river-fed margins of the basin. For effects
further into the open ocean, processes are needed that extensively redistribute
the nutrients.
The ocean has an array of magnificent tricks up
its sleeve with exactly that potential, and which involve the deep-water
nutrient reservoir. Even in a basin like the Mediterranean, with a very
active deep-sea ventilation, the intermediate and deep waters accumulate
excess nutrients due to respiration. Because the deep reservoir has an
enormous volume, even moderately enhanced nutrient levels give it a dramatic
long-term fertilisation potential (note
3). To unleash this potential, the deep nutrients need to be made
available for use in photosynthesis. In other words, the deep reservoir
needs to be brought into the zone of light penetration the photic layer,
defined as the depth of 1% light penetration, and reaching as deep as 120
m in the Mediterranean. The result of such a fertilisation process would
be a long-term sustained increase in the phytoplankton production. Depending
on where the deep reservoir is brought up to shallower depths, the fertilisation
may occur anywhere in the basin, not just around the coasts.
In the open ocean, upwelling areas are places where subsurface
water is pumped all the way to the surface by the prevailing winds. These
regions are consequently characterised by enormous production levels that
support important fisheries-resources for example the anchovy fisheries
off the coast of Peru. Upwelling is not widely developed in the Mediterranean.
A more subtle process, however, does operate in the basin. It involves
shallowing but not surfacing of the deep nutrient reservoir. In winter,
intensified surface currents pump the intermediate water from about 200
m up to about 100 m depth in the Gulf of Lions, causing a fertilisation
at the base of the photic layer. Similar subsurface fertilisation processes
are known from other places in the world ocean. Whenever this happens,
a bloom develops of a specific association of phytoplankton, which we call
the Deep Chlorophyll Maximum (DCM) association.
Based on specialisations regarding a wide variety of parameters,
the next level of the food web the secondary producers will also reflect
specific environmental conditions. To continue our DCM example, those settings
are found to sustain a specific, recognisable association of secondary
as well as primary producers. The development of such a particular association
is driven by complex interplays of parameters such as dietary preferences,
living and reproduction depth or temperature preferences, capacity to compete
for resources, and reproductive strategies to cope with predation pressure.
As a consequence, different DCM settings are never characterised by exactly
the same association of secondary producers, but distinct common aspects
may be expected.
Of course, specialisation to specific types of feeding
grounds also takes place higher up in the food web (on higher trophic
levels), but further discussion falls outside the scope of this book.
Much of the research into causes and mechanisms of the Mediterranean anoxic
events concentrates on phytoplankton and zooplankton (ie., animal plankton),
and hence to a large extent concerns primary and secondary producers. The
study of these microfossils is called micropalaeontology. The emphasis
on micropalaeontology in the Mediterranean studies did not develop by chance,
but is based on a number of sound arguments.
First, many of the phytoplankton and zooplankton species
form skeletal parts that fossilise well. Second, these skeletal parts have
different shapes and sizes for the many different species, which helps
us assess how the associations have changed through time. Third, many of
the species are highly specialised to specific conditions, facilitating
the reconstruction of past oceanic conditions. Fourth, phytoplanton and
zooplankton organisms are very small (mostly smaller than 1 mm, but occasionally
up to about 5 mm), and they occur in great numbers. This ensures that there
virtually always is enough material to perform statistically sound quantitative
analyses of species abundance, diversity, dominance, etc. even when working
with small samples from expensive sea-floor coring. Finally, the fossilised
skeletal parts of phytoplankton and, especially, zooplankton species are
excellently suitable for a variety of chemical analyses that add further
vital information to the reconstructions.
To Chapter 3
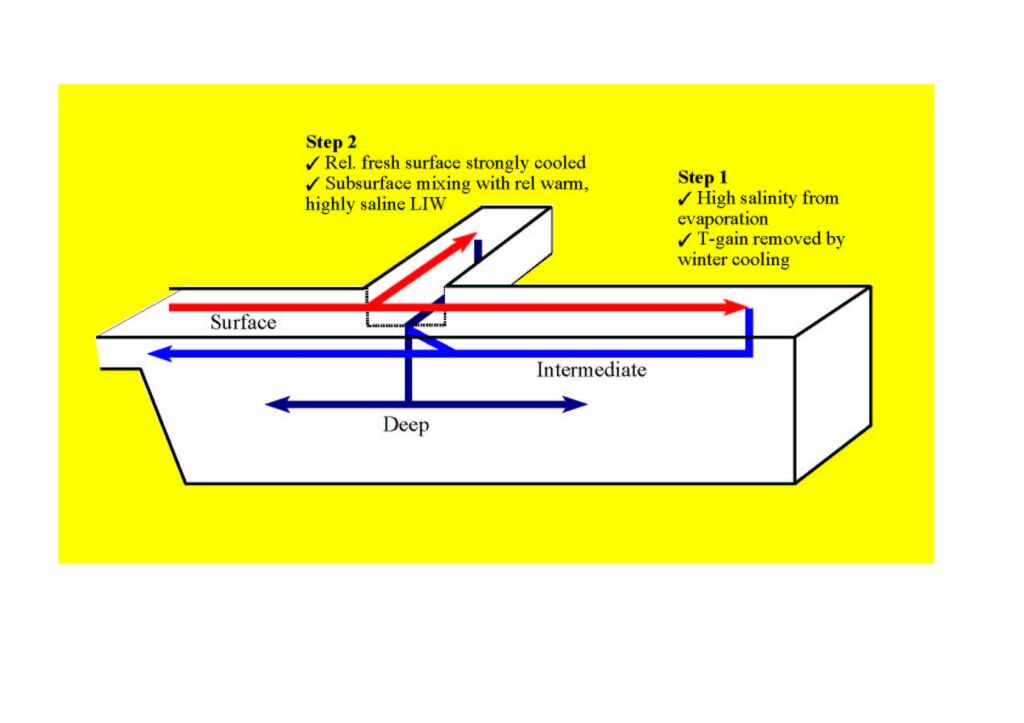 Figure 2. Highly schematic representation of the two-stage deep ventilation
engine. Inflowing surface water is transformed into intermediate water
by a net salinity increase. The salt in the intermediate layer preconditions
the second stage, whose direct forcing is essentially thermal. The endresult
is the formation of deep water layer that is less saline, but colder, than
the intermediate water. Click on thumbnail for full-sized figure.
Figure 2. Highly schematic representation of the two-stage deep ventilation
engine. Inflowing surface water is transformed into intermediate water
by a net salinity increase. The salt in the intermediate layer preconditions
the second stage, whose direct forcing is essentially thermal. The endresult
is the formation of deep water layer that is less saline, but colder, than
the intermediate water. Click on thumbnail for full-sized figure.