4. Circulation in Crisis
Global long-term monsoonal maxima develop roughly
every 21,000 years, driven by astronomical variations that form the subject
of the next chapter. The most recent monsoonal maximum began to build up
from about 11,000 years BP over North Africa, and culminated between 9,500
and 6,000 years BP. During that time, the Sahara Desert became strongly
reduced in size, by encroachment of its southernmost margin towards the
north. Saharan art by prehistoric man reflects the presence of diverse
wildlife that included giraffe, elephant, rhinoceros, hippopotamus, antelope
and ostrich, and also shows abundant humans following pastoral lifestyles.
In many places within what is today the Sahara Desert,
great lakes developed, as witnessed by their characteristic sedimentary
deposits and fossil shorelines. The locations of these lakes today present
themselves as virtually or completely dry depressions. Impressive fossil
river systems dating from the last and previous monsoonal maxima have been
found hidden at shallow depth beneath the shifting sands. Several of these
systems have been discovered by fieldwork, and others by shallow-penetrating
radar imagery from space. Some of these rivers drained eastward into the
Nile, while others flowed northward and discharged directly into the Mediterranean.
The northward shift of the Sahelian vegetation boundary that determines
the southern margin of the Sahara Desert reduced the modern South-North
extent of the desert to half its modern dimension, or less.
Aquifers (natural groundwater reservoirs) beneath the
present-day Sahara contain fossil water from the last monsoonal maximum,
with stable isotopic signatures that positively identify the South Atlantic
as its source region. Evaporation from that ocean rained out as it was
transported over the African continent by the African monsoon. The progressive
rain-out in the east-northeastward direction left a characteristic distribution
pattern in the stable isotope ratios of oxygen and hydrogen that allows
us to identify the transport direction and therefore the initial source
region of the moisture.
The African monsoon penetrated so far to the east-northeast
that it affected the Ethiopian highlands, fuelling a more vigorous seasonal
flow of the Blue Nile. The White Nile drains equatorial regions, where
rainfall is high even today, and we have little insight into its variations
through time. In any case, the Nile River formed the main pathway by which
the expanded monsoon affected the Mediterranean Sea, in particular the
easternmost sector. Rough estimates suggest 3 to 4 times stronger discharge
rates for the Nile than today, where today refers to historical values
before the completion of the Aswan dam. At the height of the monsoonal
maximum there likely were other river systems draining the African margin,
further enhancing the freshwater input into the Mediterranean Sea.
Because fresh rain and river water has low (18O
depleted) oxygen isotope ratios, it causes a measurable anomaly in sea
surface waters. Such freshwater influences are concentrated near
the surface because the low-density fresh water tends to float on the higher-density
(salty) marine water, while gradually mixing through the surface layer
by the action of wind and waves. Freshwater-induced isotope depletions
are clearly reflected in the carbonate shells of planktonic foraminifera
that live near the very surface. Statistically, the oxygen isotope depletion
that characterises the monsoonal maximum in eastern Mediterranean sedimentary
sequences gets more pronounced from West to East, suggesting a dominant
impact of the enhanced Nile outflow in the east. Other major river outflows
are identifiable as well, however, when individual isotope values are investigated
in detail.
The increase in climatic humidity and river discharges
during the monsoonal maximum was not restricted to the African margin.
Isotope data from limestone cave deposits in Israel also indicate a significantly
more humid climate at that time. In addition, there is convincing evidence
of substantially more humid climate conditions all along the European/Near
Eastern margin of the Mediterranean. Marine isotope evidence and sediment
accumulation data reflect enhanced discharge from European rivers. Pollen
data offers a further intriguing insight into the nature of the seasonal
precipitation cycle in the northern borderlands; there was not only higher
precipitation in winter, but also in summer. The Mediterranean vegetation
showed an abundance of deciduous oak between about 9,500 and 6,000 years
ago. This species requires moisture during its growth season, which is
when the tree is in foliage, through the summer. The abundance of such
oaks along the European and Near Eastern margin of the Mediterranean therefore
testifies of enhanced summer precipitation. This observation is in remarkable
contrast with the wet-winter and dry-summer climate of today, which is
termed a Mediterranean climate.
The cause of the monsoonal intensification and its impact
on precipitation over Africa is quite well understood in broad detail (next
chapter), and the bulk of the moisture transported into the Mediterranean
via this mechanism would have originated from somewhere else the South
Atlantic. The enhanced input of externally derived fresh water by the monsoonal
intensification reduced the Mediterranean excess of evaporation over total
freshwater input (excess evaporation) (note
4).
Conversely, the origin of the increased humidity over
the basins northern borderlands remains elusive, and it remains to be
seen whether this moisture was not merely a recycling of Mediterranean
water. There are three lines of thought on this issue. Firstly, microfossil
evidence from the equatorial Atlantic suggests that enhanced oceanic heat
transport occurred at this time from South to North across the equator.
This observation has fuelled speculations that Atlantic depression activity
may have been intensified as a consequence, which would bring external
moisture into the Mediterranean region. Secondly, it has been proposed
from observational evidence and computer-based climate models that the
main westerlies depression track over the Atlantic today roughly from
New York to Ireland was displaced towards the North at the European side,
reaching 65-70ºN latitude. In such a configuration, a secondary depression
track may develop over the northern Mediterranean margin. The result would
be increased external (North Atlantic) moisture supply at least into the
western Mediterranean basin, and a likely transfer of moisture from the
western to the eastern Mediterranean by way of local Mediterranean depressions.
These form in the Gulf of Genoa and track across Italy through the Plain
of Lombardy to the North Adriatic, before continuing in an east-southeastward
direction. Thirdly, the generally warmer summer conditions during the monsoonal
maximum a time when the seasonal contrast on the Northern Hemisphere
was enhanced relative today (next chapter) may have caused increased
cycling of local Mediterranean-sourced moisture over the basins borderlands.
In that case, virtually no external moisture supply would be involved,
so that the net effect on excess evaporation would be negligible.
The uncertainties about the origin and distribution mechanisms
of the northern moisture imply that despite having a distinct impact
on vegetation, river flow rates, and other humidity indicators it did
not neccessarily affect the Mediterranean evaporation-precipitation balance.
Much research is devoted to the development of a better understanding of
the origin and distribution processes of the observed humidity over the
northern borderlands. Whether due to discharge from the African margin
alone, or due to increased discharge from the African and European margins,
however, it is safe to state that the Mediterranean excess of evaporation
over total freshwater input during the apex of the monsoonal maximum was
considerably lower than today. Quantitatively, the various reconstructions
suggest quite a range of possible values, but all agree that there was
at least a 30% reduction of excess evaporation relative to the present.
* * *
The discussion of increased freshwater delivery into the
Mediterranean in terms of a reduction of excess evaporation may seem
somewhat abstract, but in fact is crucial for a further assessment of the
consequences for the Mediterraneans density structure. First, however,
we need to briefly consider the background against which the impact of
the monsoonal maximum should be evaluated. Oxygen isotope studies demonstrate
that, prior to the monsoonal maximum, the Mediterranean was a salty sea
with salinities equal to present-day values, or somewhat higher. Various
proxies indicate that average temperatures at that time were slightly lower
than today, while active intermediate and deep-water circulation ensured
that the properties were well distributed throughout the basin. High salinities
and low temperatures meant that the basin was filled with high-density
waters.
A decrease in the excess of evaporation over freshwater
input would cause a reduction in the gradient of sea surface salinities
from West to East. As freshwater input progressively increased following
the onset of the monsoonal maximum, therefore, the overall surface salinities
were effectively lowered. The geographic position of the Nile ouflow determined
that this change was especially pronounced in the easternmost sector of
the basin. With decreasing salinities, stronger cooling of surface waters
would have been needed to achieve sufficiently high densities for displacement
of previously formed salty intermediate waters. Unfortunately, temperatures
were going up, not down, so that the salt-driven first stage of the basins
subsurface ventilation inevitably began to falter.
The first stage preconditions the second stage by supplying
it with sufficient salt. Disruption of the first stage therefore spelled
disaster for the functioning of the second stage of deep ventilation, which
today affects the deep-sea below 600 m. Regarding the eastern Mediterranean
basin, polar/continental air outbreaks from the North continued to cause
strong winter cooling in the Adriatic and Aegean Seas. In other words,
the thermal forcing that today identifies these regions as sites of subsurface/deep
water formation remained in existence. The disruption of salty intermediate
water formation, however, had removed the salinity preconditioning. As
a consequence, newly forming deep waters did not achieve sufficient density
to displace the resident deep waters formed before the onset of the monsoonal
maximum. Instead, the newly formed subsurface waters settled above the
old deep waters.
So, in summary, what did the subsurface ventilation structure
look like during the monsoonal maximum? The reduction of excess evaporation
inhibited the salinity-driven formation of Levantine Intermediate Water
in the Cyprus-Rhodes area. In the absence of this salt supply, new deep-water
formation was only thermally driven. Because of the reduced salinity/density
in newly formed deep water, this water failed to sink to great depths,
and instead settled above the old salty deep waters that it could not
displace. Nowhere in the basin were high-enough densities attained to displace
the salty old deep waters. As a result, the eastern Mediterranean became
unventilated (stagnant) from about 300/400 m depth to the bottom.
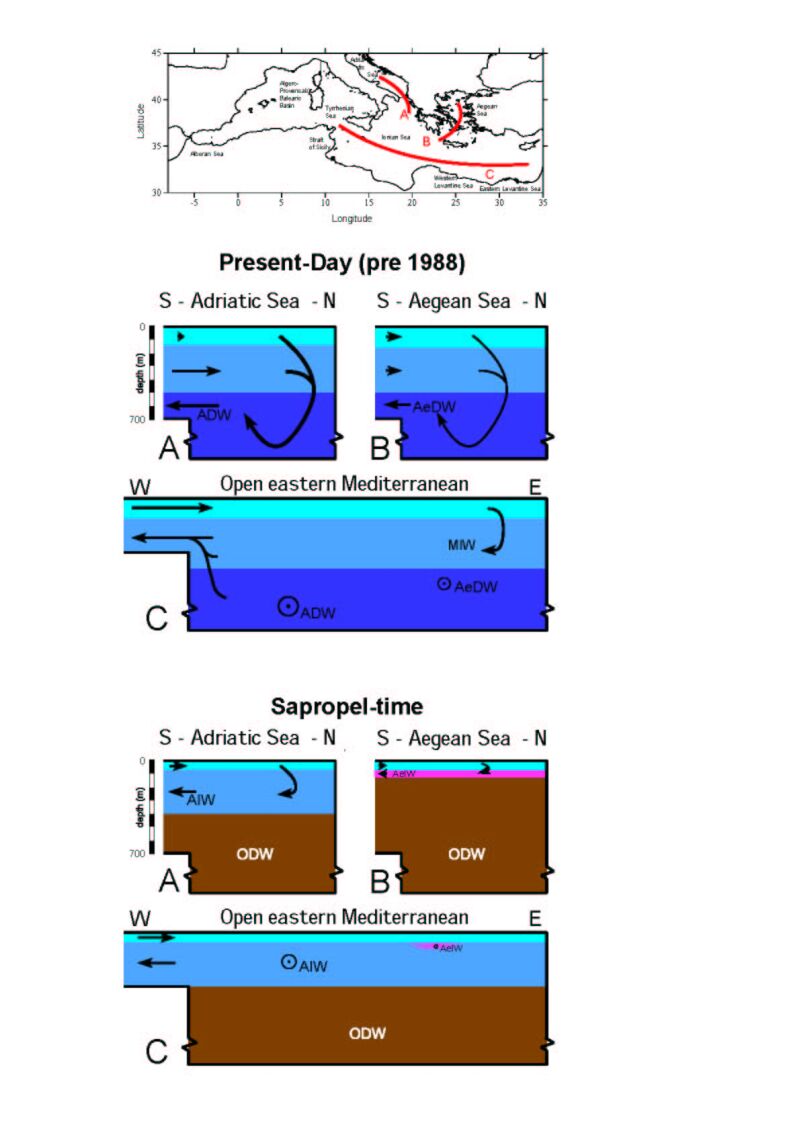
Figure 3. Schematic presentation
of the changes in subsurface circualtion patterns between the present day
and times of sapropel formation. The three profiles presented summarise
information obtained from analytical and modelling studies from North to
South through the Adriatic and Aegean basins, and from West (Strait of
Sicily) to East (near Cyprus) through the open eastern Mediterranean. MIW
stands for Mediterranean Intermediate Water; ADW for Adriatic Deep water;
AeDW for Aegean Deep water; AIW for Adriatic Intermediate Water; AeIW for
Aegean intermediate Water; ODW for Old (isolated) Deep Water. Click on
thumbnail for full-sized jpeg (or here
for a pdf).
* * *
As discussed in section 2.1., deep water ventilation constitutes
the only means of replenishing the deep sea with oxygen, while consumption
of oxygen in the deep sea is regulated by the decomposition (respiration)
of organic matter. As long as productivity continues in the surface layers,
there will be a steady supply of dead organic matter that sinks into the
deep sea, where it is subject to respiration. The demand for deep-sea oxygen
to sustain this respiration is technically known as Biological Oxygen
Demand (BOD). The disruption of deep-water ventilation below 300/400 meters
in the eastern Mediterranean, therefore, imperiled the oxygenation of the
deep sea.
Even with the fairly low present-day levels of production
of organic matter in the surface layers of the eastern Mediterranean, there
is a notable BOD in the deep sea. In fact the BOD in the Mediterranean
is surprisingly high given the low rate of production, when compared with
other places in the world ocean. This is thought to be due to the high
deep-sea temperatures in the Mediterranean (around 13ºC), compared
with those in the open ocean (typically 3ºC or less). Metabolism speeds
up with increasing temperature, and a higher metabolic rate necessitates
more oxygen intake by the deep-sea organisms. Following a cessation of
deep ventilation, all oxygen in the Mediterranean deep sea would be consumed
in roughly 500 to 1,000 years at modern BOD values.
During the monsoonal maximum, eastern Mediterranean productivity
and therefore BOD in the deep sea were higher than today, shortening the
time needed for depletion of all oxygen from the stagnant (unventilated)
old deep waters. Several proxies reflect the higher productivity in the
basin at that time (note
5), and also identify the likely mechanism by which the productivity
increase was established. Key to the discovery of this mechanism was the
observation that zooplankton and phytoplankton assemblages in the eastern
Mediterranean anoxic intervals, associated with monsoonal maxima, were
systematically different than today.
In 1989, it was noted for the first time that planktonic
foraminiferal faunas in the anoxic intervals (technically known as sapropels)
commonly contain strong abundances of species that are known to prefer
settings with a distinctly developed Deep Chlorophyll Maximum (DCM; see
section 2.3.). The study also demonstrated that these DCM faunas prevailed
on a basin-wide scale throughout the eastern Mediterranean, suggesting
a fundamental change in the mode of productivity and the underlying water-column
structure. The DCM hypothesis was later corroborated by a study of coccolith
abundances in several sapropels, including the most recent one of 9,500
to 6,000 years BP. Further confirmation came from a study of the unusual
preservation of diatom floras in a sapropel that dates from 125,000 years
BP.
A deep chlorophyll maximum is a phytoplankton productivity
maximum near the base of the photic layer ie. in the shade zone with
little over 1% light penetration. To establish a DCM, a combination is
needed of sufficient (if low) light intensity from above and a steady
supply of nutrients. The facts that such conditions developed on a basin-wide
scale, and consistently remained present for a few thousand years, suggest
that the nutrient supply was somehow related to the subsurface nutrient
reservoir (note 6).
Therefore, the most likely mechanism of hydrographic change had to be identified
that would make the subsurface nutrient reservoir available at the base
of the photic layer (100-120 meters depth). It was proposed that changes
in the density structure of the basin, due to the enhanced freshwater input,
raised the top of the intermediate water from well below the base of the
photic layer as today to a shallower depth within the photic layer.
The boundary between intermediate and surface waters today
resides at about 150 meters depth in the eastern Mediterranean. There is
no distinct DCM due to the fact that the subsurface nutrient reservoir
accessible in the intermediate layer remains well below the base of
the photic layer (100-120 m). The DCM hypothesis for times of sapropel
formation invokes a shallowing of that boundary to roughly half its modern
depth. The resultant overlap of the subsurface nutrient reservoir with
the lower photic zone throughout the basin was proposed to have sustained
the DCM-indicative phytoplankton and zooplankton assemblages. Simple box-model
exercises in the early 1990s confirmed that this would be a likely response
of the basins density structure to a reduction in excess evaporation.
The box-models also showed that comparable responses should be expected
due to major drops in sea level. This was interesting, since observations
in glacial maximum intervals when global sea level stood more than 100
meters below the present mark had also revealed high abundances of DCM
indicators.
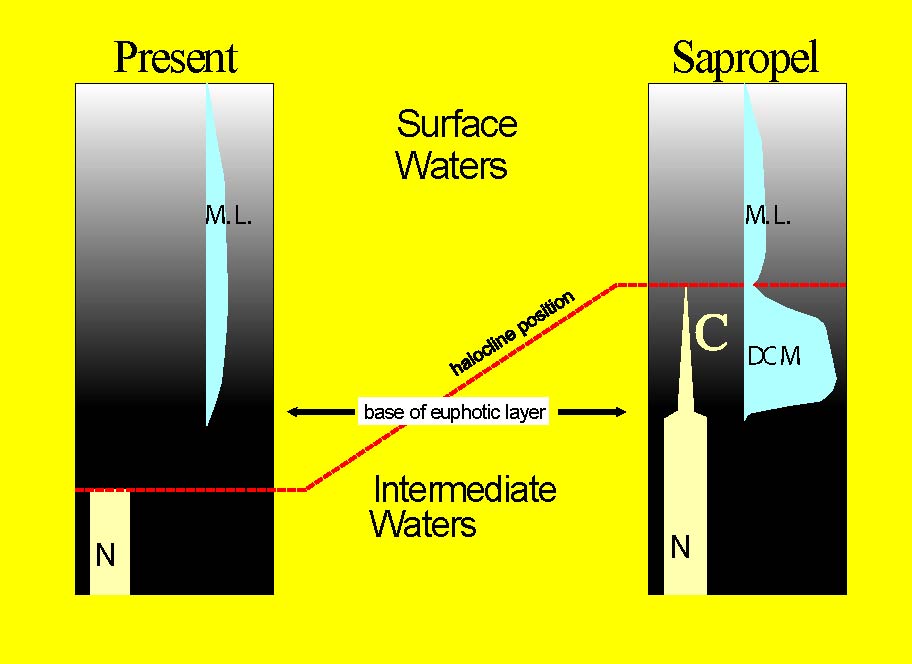
Figure 4. Difference in the
depth of the surface to intermediate water interface between the present
and times of sapropel deposition, relative to the depth of light penetration
(base euphotic layer = 1% light intensity level). N indicates nutrients,
C indicates consumption of nutrients for photosynthesis, DCM stands for
Deep Chlorophyll Maximum association, and M.L. for Mixed Layer association.
Click on thumbnail for full-sized jpeg (or here
for a pdf).
In 1998, a computer-based numerical model was presented
to evaluate possible modes of Mediterranean circulation during periods
of sapropel deposition. A wide variety of surface salinity fields was used
to cover all the different reconstructions proposed in the literature.
For all fields, the numerical model predicted a single, robust, circulation
pattern. This pattern was found to be in close agreement with the
observations and the previous rather qualitative interpretations.
The numerical model predicted that the formation of salty
Levantine Intermediate Water (LIW) would have been disrupted during the
monsoonal maximum. In the absence of the LIWs salt supply, only thermal
forcing remained in the Adriatic Sea, driving the formation of a new intermediate
water mass that spread throughout the basin between about 80 and 350 meters
depth. Below 350 meters a stagnating body of salty old deep water remained.
Although the quoted depth range should not be taken too absolutely, it
revealed an intriguing match with the DCM hypothesis: the upper limit of
the new intermediate-water mass at 80 m would be well within the photic
zone (note 7). The
model therefore provided a strong confirmation that DCM development and
isolation of old deep water represent two distinct and verifiable responses
that are intimately related to a single forcing factor a reorganisation
in the basins hydrographic structure driven by a reduction of excess evaporation.
* * *
Unventilated and suffering the enhanced biological oxygen
demand caused by a basin-wide increase in productivity (and hence sinking
organic matter), the old deep water below about 350 meters depth inevitably
became anoxic. The complete absence of fossils of benthic organisms, and
preservation of original sedimentary laminae, demonstrate that the anoxicity
wiped out the deep-sea ecosystem. Organic matter sinking through the anoxic
water column was not as effectively broken down as it would be under fully
oxygenated conditions, and an excess of organic matter started remained
preserved in the sediments. Delicate organic fossils were preserved, testifying
to the less efficient degradation of organic matter in the absence of oxygen.
A sapropel was being deposited.
To Chapter 5